WHO announced the official disease name as COVID-19 caused by the newly named virus SARS-CoV-2 to replace the former “2019-nCoV” (International Committee on Taxonomy of Viruses).
Although people don’t like to say it, COVID-19 is a pandemic. It’s been shown to appear on pockets and demographics that do not seem to have any direct contact/relation to China.
Origins & Genome
2019-nCoV was first found & recognized in Wuhan, China.
Genetically closest to SARS-CoV (shared 79.5% similarity) in the Betacoronaviruses (Nidovirales, Coronaviridae, Coronavirinae, Betacoronavirus, Sarbecovirus), and also shares similiarities with bat SARS-like variants, possibly BatCoV RaTG13 (96.2%).
Virus was thought to have crossed over to humans from bats.
The report trying to link HIV to 2019-nCoV was erroneous.
Reports of 2019-nCoV originating from snakes were erroneous.
Produces acute respiratory illnesses, fever, cough, difficulty breathing/shortness of breath.
2019-nCoV targets the angiotensin converting enzyme 2 (ACE2) receptor.
Incubation ranges between 2-14 days. Some say up to 20 days.
Use standard airborne disease precautions.
The severity of symptoms is variable.
Babies and children have mostly been spared.
There have been some cases of people being re-infected, but that is not too clear.
Testing has been somewhat unclear at this point.
Transmission
Human-to-human spread up to approximately 6 feet distance (CDC, 02.05.2020) via airborne droplets from in infected person (e.g. sneezing, coughing, etc.).
High sputum viral load.
The virus may persist on surfaces but only for a relatively “short time”.
The virus has been found in feces, but the main transmission method is still in droplets–airborne.
People who carry the virus but don’t show symptoms can still spread the disease, asymptomatic spreading.
Hand hygiene is important. As taught in nursing school: use warm water; use antibacterial soap to vigorously scrub front-back-nails of hand for at the very least 20 secs. DO NOT TOUCH the faucet or anything else. Dry your hands and use the paper towel to shut off the water. If your paper towel dispenser is not automatic/motion-controlled, then before you wash your hands pull some paper towel down so that you can avoid touching the dispenser after your hand wash. https://www.cdc.gov/handwashing/when-how-handwashing.html
Centers for Disease Control (CDC), & National Center for Immunization and Respiratory Diseases (NCIRD), Division of Viral Diseases. (2020, January 31). Transmission of Novel Coronavirus (2019-nCoV). Retrieved February 5, 2020, from https://www.cdc.gov/coronavirus/2019-ncov/about/transmission.html
Cohen, J. (2020, January 31). Mining coronavirus genomes for clues to the outbreak’s origins. Retrieved February 5, 2020, from https://www.sciencemag.org/news/2020/01/mining-coronavirus-genomes-clues-outbreak-s-origins
European Centre for Disease Prevention and Control (ECDC). (2020, January 31). Disease background of 2019-nCoV. Retrieved from https://www.ecdc.europa.eu/en/2019-ncov-background-disease
Greens, K. (2020, February 4). Report of asymptomatic transmission of 2019-nCoV inaccurate. Retrieved from https://www.the-scientist.com/news-opinion/report-of-asymptomatic-transmission-of-2019-ncov-inaccurate-67060
Kupferschmidt, K. (2020, February 3). Study claiming new coronavirus can be transmitted by people without symptoms was flawed. Retrieved from https://www.sciencemag.org/news/2020/02/paper-non-symptomatic-patient-transmitting-coronavirus-wrong
The New England Journal of Medicine (NEJM). (2020, January 30). Transmission of 2019-nCoV infection from an asymptomatic contact in Germany. Retrieved from https://www.nejm.org/doi/full/10.1056/NEJMc2001468
Paraskevis, D., Kostaki, E. G., Magiorkinis, G., Panayiotakopoulos, G., Sourvinos, G., & Tsiodras, S. (2020). Full-genome evolutionary analysis of the novel corona virus (2019-nCoV) rejects the hypothesis of emergence as a result of a recent recombination event. Infection, Genetics and Evolution, 104212.
Robertson, D. L. (2020, January 26). nCoV’s relationship to bat coronaviruses & recombination signals (no snakes) – no evidence the 2019-nCoV lineage is recombinant. Retrieved February 5, 2020, from https://virological.org/t/ncovs-relationship-to-bat-coronaviruses-recombination-signals-no-snakes-no-evidence-the-2019-ncov-lineage-is-recombinant/331
Coronaviruses are responsible for one-third of the common colds and respiratory illnesses (ranging from mild to severe) among humans like upper respiratory tract infections (URTIs) and lower respiratory tract infections (LRTIs bronchitis, pneumonia, severe acute respiratory syndrome) (Burrell, Howard, & Murphy, 2016; Li et al., 2019; Shoeman & Fielding, 2019). These viruses are higly adaptable and prone to mutation (selective pressure, environment).
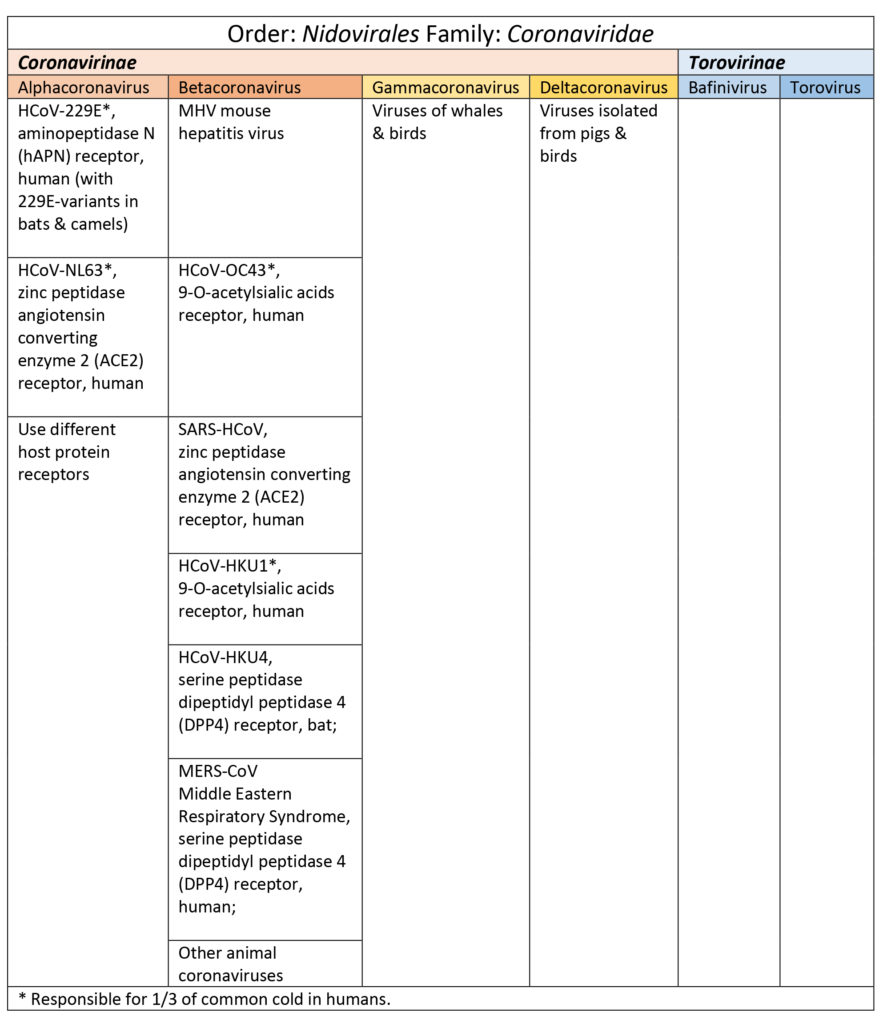
Coronaviruses belong to the order Nidovirales and family Coronaviridae. There are two subfamiles: Coronavirinae (spherical) and Torovirinae (disc, bean or rod-shaped and pertain mostly to horses, cattle, pigs, cats, and goats illnesses). Coronaviruses get their name from their unique from their “fringe-like” envelope with glycoproteins sticking out looking like a crown or crown-of-thorns (Burrell et al., 2016; Shoeman & Fielding, 2019).
Coronavirinae are subdivided into four genus categories: alpha, beta, gamma (whales, birds), & delta (pigs & birds). Coronaviruses in the alpha and beta categories viruses that have affected humans are HCoV-229E, HCoV-NL63, HCoV-OC43, SARS-HCoV, HCoV-HKU1, and MERS-CoV (Burrell et al., 2016; Shoeman & Fielding, 2019). See Table 1. SARS, SARS-like, and MERS-CoV share a good percentage of their genome with bat coronaviruses (Burrell et al., 2016; Shoeman & Fielding, 2019).
Table 1
Structural Proteins
Coronaviruses have 4 key structural proteins S, M, N, and E. S (spike) protein is a large glycoprotein responsible for forming the peplomers (glycoprotein spikes which will selectively bind to certain receptors) of the envelope. M is a transmembrane glycoprotein. N is a phosphorylated nucleocapsid. Additionally, E is a envelope protein and some species of betacoronaviruses also have protein HE (hemagglutination and esterase functions) (Burrell et al., 2016; Shoeman & Fielding, 2019; Tok & Tatar, 2017). Interestingly, not all four key structural proteins are required for producing a viable virion which suggests there may be some redundancy in the genome (Shoeman & Fielding, 2019). The S, M, N, and E proteins also participate in viral replication.
Viral Genome
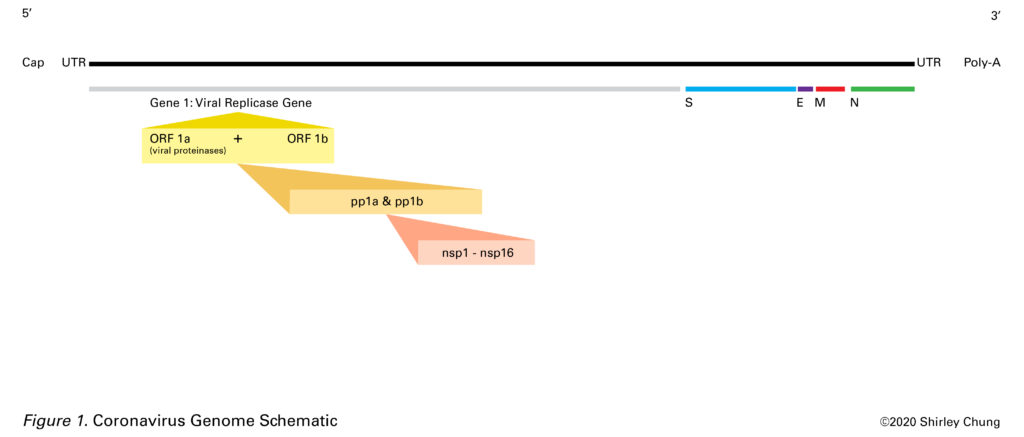
Coronaviruses have a single strand positive-sense RNA (+ssRNA) 26-32 kilobases (kb) long (Burrell et al., 2016; Shoeman & Fielding, 2019). This +ssRNA is of the same sense as host mRNA which allows the virus to harness the host cell’s machinery. The 5′ end (has untranslated sequence, UTR, of 65-78 nucleotides or leader RNA) is capped and the 3′ end has 200-300 UTR nucleotide sequence plus the poly-A tail (Burrell et al., 2016; Shoeman & Fielding, 2019). The 5′ cap is a way for the virus to disguise its genetic material to make it similar to host mRNA in order to use the host’s “machinery” (Picard-Jean, 2013).Nested mRNAs all share the same 5′ end sequence (Burrell et al., 2016; Shoeman & Fielding, 2019). Untranslated regions (UTRs) help regulate replication and transcription.
There are 7-14 open reading frames (ORFs) which are a sequence of codons that begin with a start codon and end with a stop codon (Burrell et al., 2016). These ORFs are interspersed within the genome.
Starting at the 5′ end (plus UTR), gene 1 (20-22 kilobases long) is about two-thirds of the entire RNA length. The spike (S) protein gene comes next followed by the E (envelope) protein gene, M (transmembrane) protein gene, the N (nucleocapsid) gene, another UTR sequence plus poly-A tail at the 3′ end (Burrell et al., 2016; Li, 2016; Shoeman & Fielding, 2019). Other ORF proteins, the HE glycoprotein gene, and accessory protein genes are interspersed amongst the aforementioned major segments (Burrell et al., 2016; Shoeman & Fielding, 2019; Tok & Tatar, 2017). Burrell et al. (2016) noted that ORFs may code for accessory proteins that may account for variability and novel species. The SARS-CoV RNA contains smaller ORFs at the 3′ end that are unique to SARS-CoV (Burrell et al., 2016). Accessory proteins may also contribute to viral hardiness.
The Viral Replicase Gene (aka Gene 1)
Gene 1 contains overlapping ORFs 1a and 1b (Burrell et al., 2016). Together, ORFs 1a and 1b constitute the viral replicase gene (gene 1) and encode for two large polyproteins (pp1a and pp1b, proteins that are covalently conjoined) (Nakagawa, Lokugamage, & Makino, 2016). In general, replicase is an enzyme that catalyzes the production of a complementary RNA strand based off of an RNA template. Proteases help cleave out 15-16 nonstructural proteins (nsp1-nsp16) from the two polyproteins to be used in the viral lifecycle (Nakagawa et al., 2016). Nsp 2-nsp16 are critical to viral RNA transcription and replication (Nakagawa et al., 2016).
The S (Spike) Protein
The S glycoprotein is important as it plays a major role in the first step towards creating viral progeny. The spike determines if a virus can fuse with a host and enter. Successful fusion is receptor-dependent. S is a clove-shaped homotrimer (protein made of 3 identical subunits) (Li, 2016). The S protein is also responsible for possible multiple cell fusions between the infected and adjacent cells, syncytia (Shoeman & Fielding, 2019).
There are three classes of fusion proteins based on the structure of their fusion subunits: class I, mostly alpha-helical; class II mostly beta-structures; class III both alpha-helical and beta-structures (Harrison, 2012; White & Whittaker, 2016). Coronaviruses S protein is class I (White & Whittaker, 2016). Other class I viruses include some forms of influenza, human immunodeficiency virus (HIV), and Ebola (Li, 2017).
The spike has regions: large ectodomain (the part that sticks out into the extracellular space where upon contact may initiate signal transduction); single-pass (crosses membrane only once) transmembrane anchor (TM1); and intracellular tail (Li, 2016). The ectodomain contains S1 (trimeric head) and S2 (trimeric stalk) (Li, 2017).
For many coronaviruses, host cell enzymes (proteases) break down S into subunits S1 and S2 (Tortorici et al., 2019). Before fusion occurs, S1 and S2 are non-covalently held together. The N-terminal S1 subunit is made of four areas A, B, C, and D. A and B are the receptor-binding areas in coronaviruses (Tortorici et al., 2019). The C-terminal S2 subunit is where the “fusion machinery” is located allowing for S2 to fuse the viral and host membranes (Li, 2016; Tortorici et al., 2019). Upon entry into the host, S2 is cleaved again at S2′ (this happens for all coronaviruses) and scientists think it may be a step that activates the protein for membrane fusion as it produces irreversible conformational changes (Tortorici et al., 2019).
Post-fusion, the pre-fusion three heads of S2 become a six-bundled helical structure with fusion peptides sticking out (Li, 2017).
The N (Nucleocapsid) Protein
N is the only protein responsible for phosphorylated helical nucleocapsid (“shell” enclosing the genome) (Shoeman & Fielding, 2019). The N protein is located near the endoplasmic reticulum (ER) and Golgi complex area. Expression of the N gene was found to coincide with the increased production of virus-like particles (VLPs), and scientists believe that N may contribute towards producing whole and viable virions (Shoeman & Fielding, 2019).
The M (Transmembrane) Protein
The M glycoprotein has three transmembrane areas and influences the shape of the envelope (Shoeman & Fielding, 2019; Tok & Tater, 2017). In the Golgi complex, a carbohydrate unit is attached to the M protein (glycosylation which is important for protein folding) which contributes to the protein’s ability to bind at receptor sites (Tok & Tatar, 2017).
M may also have an “organizational” role as it interacts with the other structural proteins (Shoeman & Fielding, 2019). M seemed to interact with S protein to “corral” or aggregate S in the ER-Golgi intermediate compartment (ERGIC, mediate transport between the ER and Golgi complex) thus allowing for packaging into new virions (Shoeman & Fielding, 2019). M interacting with N increased nucleocapsid stability (Shoeman & Fielding, 2019). M and E protein interaction contributed to the maintenance of the envelope, the production and release of VLPs (Shoeman & Fielding, 2019).
The E (Envelope) Protein
E proteins (short integral membrane proteins 76-109 amino acids long) are the smallest structural proteins and the least understood. At the hydrophilic N-terminal, E has 7-12 amino acids followed by a transmembrane domain (TMD) 25 amino acids long, and ending with a long hydrophilic C-terminal (Shoeman & Fielding). Surprisingly, E protein was not found to be abundant embedded in the viral envelope which suggested that E may serve in other capacities around the ER, Golgi, and ERGIC locations (Shoeman & Fielding, 2019). E proliferation was found inside the host (near ER, Golgi complex, ERGIC) during viral replication, and the lack of E protein seemed to have deleterious effects on viral progeny (Shoeman & Fielding, 2019).
An unique feature of coronaviruses is that they bud into the ERGIC where they get packaged into their envelope (Shoeman & Fielding, 2019). In contrast, other viruses bud only from the inside face of the plasma membrane. E protein was found to play a role in scission to complete the release of viral progeny.
Transmembrane areas (TMDs) are important for the viral ability to fuse with a host (Webb, Smith, Fried, & Dutch, 2018). Viroporins are a type of transmembrane protein. Viroporins are small viral pH-sensitive pore-inducing proteins (60-120 amino acids long, mostly hydrophobic) that can create passageways (hydrophilic pores selective for H+, K+, Na+, Ca2+ but also able to transport anions) for the virus to utilize during different reproductive activities of its life cycle (Schoeman & Fielding, 2019). The lack of viroporins was found to mediate viral pathogenicity. Targeting a virus’s viroporin may be one way to interrupt it’s activities.
The HE (Hemagglutination and Esterase) Protein
The HE protein (found in some betacoronaviruses) is associated with the viral envelope. HE acts like hemagglutinin (HA) and binds to sialic acid (Sia) which is a derivative of neuraminic acids, 9-carbon backbone acidic sugars (N-acetylneuraminic acid) (Chemspider, n.d.; Langereis, Zeng, Heesters, Huizinga, & De Groot, 2012; Mora-Díaz, Piñeyro, Houston, Zimmerman, & Giménez-Lirola, 2019; Zanin, Baviskar, Webster, & Webby, 2016). Sia is sometimes used synonymously as N-acetylneuraminic acid. Sia is important in maintaining a mucous defense. HA binds to Sia of mucins and disrupts this mucous defense system (Mora-Díaz et al., 2019; Zanin et al., 2016).
Clinical
Coronaviruses favor and tend to first replicate in the respiratory or intestinal tracts (Burrell et al., 2016). They may be stable for some time outside of the human body on environmental/object surfaces (fomites) (Burrell et al., 2016). Coronaviruses may also be found in stools. Mouse hepatitis virus (MHV) is a type of betacoronavirus that can be found not only in the respiratory/intestinal tracts but also causing liver and central nervous system infections (Burrell et al., 2016). The neurological effects of MHV have been studied extensively due to its similarity to multiple sclerosis (MS) in humans (Burrell et al., 2016). Cross-immunity between different coronaviruses does not seem likely (Burrell et al., 2016). Coronaviruses are difficult to detect as an infected person may remain asymptomatic. One challenge in studying coronaviruses is that they are difficult to grow in vitro.
Closing
In order to take a step forward to try understand 2019-nCoV, it was necessary to take a step backward and try understand some of the general features of the coronaviruses. Moving forward, the next step is to learn about some of the unique features of 2019-nCoV. Undoubtedly, such knowledge at this nascent stage of viral outbreak remains fluid and dynamic.
References
Burrell, C. J., Howard, C. R., & Murphy, F. A. (2016). Fenner and White’s Medical Virology, 5.
Chemspider. (n.d.). N-Acetylneuraminic acid | C11H19NO9. Retrieved February 4, 2020, from https://www.chemspider.com/Chemical-Structure.10292217.html
Harrison, S., & Harvard Howard Hughes Medical Institute (HHMI). (2012a, March 24). Part 2: Viral membrane fusion [Video]. Retrieved from https://youtu.be/qcepGvFUM38
Harrison, S., & Harvard Howard Hughes Medical Institute (HHMI). (2013b, November 1). Class 1 viral fusion proteins [Video]. Retrieved from https://youtu.be/FcX9mD8rvYw
Langereis, M. A., Zeng, Q., Heesters, B., Huizinga, E. G., & De Groot, R. J. (2012). The murine coronavirus hemagglutinin-esterase receptor-binding site: a major shift in ligand specificity through modest changes in architecture. PLoS pathogens, 8(1).
Li F. (2016). Structure, Function, and Evolution of Coronavirus Spike Proteins. Annual review of virology, 3(1), 237–261. doi:10.1146/annurev-virology-110615-042301
Li, Z., Tomlinson, A. C., Wong, A. H., Zhou, D., Desforges, M., Talbot, P. J., … & Rini, J. M. (2019). The human coronavirus HCoV-229E S-protein structure and receptor binding. eLife, 8.
Mora-Díaz, J. C., Piñeyro, P. E., Houston, E., Zimmerman, J., & Giménez-Lirola, L. G. (2019). Porcine Hemagglutinating Encephalomyelitis Virus: A Review. Frontiers in veterinary science, 6.
Nakagawa, K., Lokugamage, K. G., & Makino, S. (2016). Viral and cellular mRNA translation in coronavirus-infected cells. In Advances in virus research (Vol. 96, pp. 165-192). Academic Press.
National Human Genome Research Institute (NHGRI). (n.d.). Open Reading Frame. Retrieved from https://www.genome.gov/genetics-glossary/Open-Reading-Frame
Picard-Jean, F., Tremblay-Létourneau, M., Serra, E., Dimech, C., Schulz, H., Anselin, M., … & Bisaillon, M. (2013). RNA 5′-end maturation: a crucial step in the replication of viral genomes. Current Issues in Molecular Virology: Viral Genetics and Biotechnological Applications, 27.
Schoeman, D., & Fielding, B. C. (2019). Coronavirus envelope protein: current knowledge. Virology journal, 16(1), 69.
Tok, T. T., & Tatar, G. (2017). Structures and Functions of Coronavirus Proteins: Molecular Modeling of Viral Nucleoprotein.
Tortorici, M. A., Walls, A. C., Lang, Y., Wang, C., Li, Z., Koerhuis, D., … & Veesler, D. (2019). Structural basis for human coronavirus attachment to sialic acid receptors. Nature structural & molecular biology, 26(6), 481-489.
University of Washington Health Sciences/UW Medicine. (2016, February 26). Electron microscopy captures snapshot of structure coronaviruses use to enter cells: Atomic model suggests vaccine strategies against deadly pandemic viruses such as SARS-CoV and MERS-CoV. ScienceDaily. Retrieved January 31, 2020 from www.sciencedaily.com/releases/2016/02/160226173209.htm
Webb, S. R., Smith, S. E., Fried, M. G., & Dutch, R. E. (2018). Transmembrane Domains of Highly Pathogenic Viral Fusion Proteins Exhibit Trimeric Association In Vitro. mSphere, 3(2), e00047-18.
White, J. M., & Whittaker, G. R. (2016). Fusion of enveloped viruses in endosomes. Traffic, 17(6), 593-614.
Zanin, M., Baviskar, P., Webster, R., & Webby, R. (2016). The interaction between respiratory pathogens and mucus. Cell host & microbe, 19(2), 159-168.
From our previous post on R “nought” (the basic reproduction number), R0 is a best estimation and defined as: “the average number of secondary infections generated by the first infectious individual in a population of completely susceptible individuals” (CIDD, 2014a; CIDD, 2014b).
If R0>1, then the outbreak is likely to continue; if R0<1, then the outbreak has a better potential for being contained (Delamater et al. 2019). Anticipated outbreak/epidemic size is commonly based on R0 as is the estimation of the number of people needing to be vaccinated (Delamater et al., 2019).
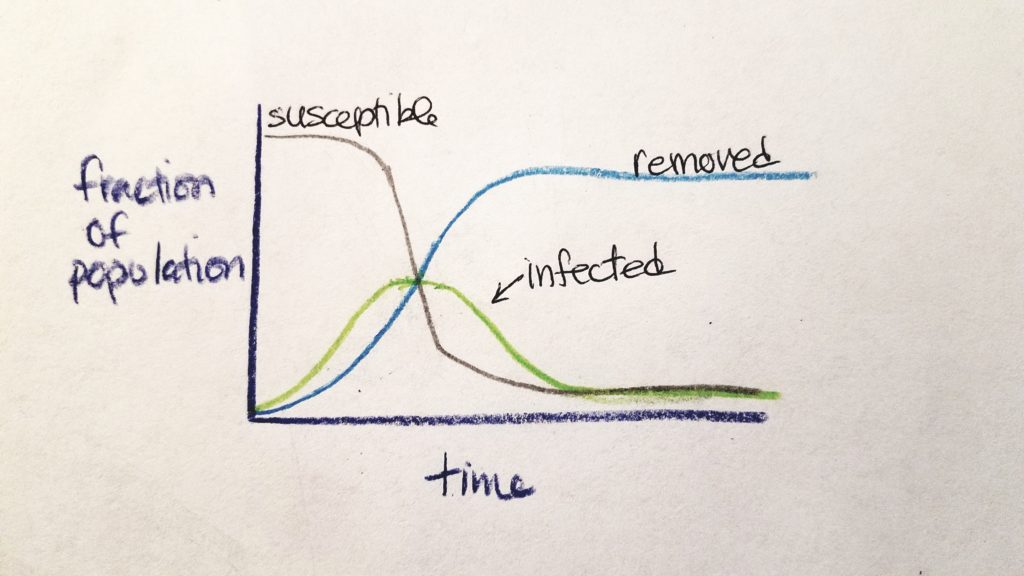
R0 values are calculated early at the onset of an outbreak. As time goes on, the nature and numbers of the outbreak change. At the beginning, most people are susceptible and the rate of potential transmission is exponentially great. As time goes on, people are either susceptible (S), infected (I), or removed from the chain of infection as non-susceptible (R) (either they are immune and regain health or they are dead) (CIDD, 2014b). The rate of susceptible people and infected people decreases as the rate of removed people increases.
This was explained by CIDD (2014b) as a “stylized” model called SIR Compartmental Framework (S=susceptible, I=infected, R=removed).
The speed of increase of infected individuals depends on R0 and the infectious period–higher R0 and shorter infectious period models a more prolific spread of the disease (CIDD, 2014b). As time goes on and assuming that more individuals are not added into the original population, the number of susceptible individuals decrease (either they died or regained their health).
The effective reproductive number (RE) is the average number of new infections later in the epidemic. RE(t) is defined as a function of time (t): the average number of secondary infections caused by a newly infected individual at time, t (Chirombo, Diggle, Terlouw, & Read, 2018).
RE is a product of R0 and the fraction of susceptible population: RE = R0 x fraction_of_susceptible_population (CIDD, 2014b). If RE < 1, the epidemic is self-limiting and said to be under control due to countermeasures (CIDD, 2014b; Chirombo et al., 2018). When the proportion of susceptible individuals is less than 1/R0, we call this “the point at which the population reaches herd immunity” (CIDD, 2014b).
Herd immunity is best defined by Wikipedia (2002) as: “a form of indirect protection from infectious disease that occurs when a large percentage of a population has become immune to an infection, thereby providing a measure of protection for individuals who are not immune”. Other definitions mention vaccination, but Wikipedia’s definition is more generalized (and most applicable) while retaining the same meaning.
As of January 26th, 2020 Zhang & Wang (2020) estimated RE for 2019-nCoV to range between 3.3-5.5 which concur with other studies (Cao et al., 2020).
Cao, Z., Zhang, Q., Lu, X., Pfeiffer, D., Jia, Z., Song, H., & Zeng, D. (2020). Estimating the effective reproduction number of the 2019-nCoV in China. medRxiv. https://doi.org/10.1101/2020.01.27.20018952
Center for Infectious Disease Dynamics (CIDD). (2014a, March 19). Week 1 Video 5: Reproductive Number [Video]. https://youtu.be/ju26rvzfFg4
Center for Infectious Disease Dynamics (CIDD). (2014b, March 19). Week 1 Video 6: Epidemic Curve [Video]. https://youtu.be/sSLfrSSmJZM
Chirombo, J., Diggle, P. J., Terlouw, D. J., & Read, J. M. (2018). Estimation of spatially varying effective reproduction numbers for infectious disease epidemics. Modelling spatial processes of infectious diseases, 39.
Delamater, P. L., Street, E. J., Leslie, T. F., Yang, Y. T., & Jacobsen, K. H. (2019). Complexity of the basic reproduction number (R0). Emerging infectious diseases, 25(1), 1.
Wikipedia. (2002, September 18). Herd immunity. https://en.wikipedia.org/wiki/Herd_immunity
Zhang, C., & Wang, M. (2020). Origin time and epidemic dynamics of the 2019 novel coronavirus. bioRxiv.
Many factors contribute to how disease is spread including (but not limited to): the type of pathogenic microbe; the lifespan of the microbe; any special features of the microbe; the disease vector (and its location, movement, lifespan, lifestyle characteristics/habits); the location/weather/natural barriers/migration patterns; the age/health/nutrition profile/lifestyle/habits of the infected person and his/her location plus the number of people that the infected person may encounter daily; the people in daily contact with the infected person and these people’s age/health/nutrition profile/lifestyle etc.
These details are elements of the epidemiologic triad (agent, host, and environmental factors) (Delamater, Street, Leslie, Yang, & Jacobsen, 2019). In epidemiology, R0 is used to estimate how infectious a pathogen might be (especially novel pathogens and potential outbreaks).
R “nought” or R0 is the basic reproduction number is defined as “the average number of secondary infections generated by the first infectious individual in a population of completely susceptible individuals” (CIDD, 2014a; CIDD, 2014b).
For example, consider person0 who is infected. How long has that person0 been infected? The longer person0 has been infected, the more opportunity there is for person0 to be in contact with more people and potentially infect them. The length of infection (L) is a variable that can increase the value of R0 (CIDD, 2014a; Cintrón-Arias, 2015).
Susceptible hosts (S) are (in this case we are talking about) other people who can potentially catch the disease from person0 (CIDD, 2014a; Cintrón-Arias, 2015 ). If person0 knows a lot of people and is in contact with a lot of people, then the number of susceptible hosts (S) is larger–increase the number of potentially infected people (R0 increases). Also, consider where person0 is living. If person0 is living in New York City where the population density is very high, S can also increase. This may continue on exponentially (CIDD, 2014a; CIDD, 2014b).
R0 also depends on transmissability–how transmissable is a pathogen. What affects transmissability? The properties of the pathogen and the population being studied affect transmissability (B, “beta”). The rate of potentially transmissable contacts (PT) and the likelihood of a successful transmission (ST, the infected person gets sick) are factors in T (CIDDa, 2014).
ST depends on the characteristics of the pathogen. Some pathogens are very hardy and prolific. For example, you can catch the pathogen just by being in the same room as the sick person. Some pathogens are very fragile and “finicky” making them more difficult to be successfully transmitted–they have many more environmental requirements in order for them to be viable.
Potentially transmissable contacts (PT) is dependent on the characteristics of the population. More susceptible people include those who are very young/elderly, people lacking good nutrition, people who already have other health problems, etc.
R0 is a function of S, L, and B. R0 may be a single value or a low-to-high range. Remember that R0 is a “best guess” for the current situation–it is not definitive. In the case of a novel pathogen, R0 must be modeled off of knowledge of past outbreaks with known pathogens. Complex math, artificial intelligence, dynamic computation, and scientists from many different fields contribute to such work.
If R0>1, then the outbreak is likely to continue; if R0<1, then the outbreak has a better potential for being contained (Delamater et al. 2019). Anticipated outbreak/epidemic size is commonly based on R0 as is the estimation of the number of people needing to be vaccinated (Delamater et al., 2019).
Using available data for 2019-nCoV through January 21st, 2020, Read, Bridgen, Cummings, Ho, and Jacobsen (2019) estimated R0 (model based on human-human transmission, omitting zoonotic transmission) to be 3.8 (95% confidence interval, 3.6 and 4.0) implicating that 72-75% of transmissions must be prevented/controlled in order to stop the increase. Read et al. (2019) also estimated that only 5.1% (95% confidence interval, 4.8-5.5) of the infections in Wuhan have actually been identified/confirmed. It was very likely that the information and statistics released to the media (especially from the Chinese government) were under-reported. Predicted numbers were closer to 191,529 infections by February 4th, 2020 (Read et al., 2019). From rough estimates, R0 for 2019-nCoV are significantly greater than the R0 for MERS-CoV, but closer to the R0 for SARS (Read et al., 2019).
Using Virus Host Prediction (VHP), Zhu et al. (2020) found the infectivity pattern of 2019-nCoV to be more similar to Bat SARS-like coronavirus and mink coronavirus. The Huanan seafood and wet market included sales of produce, mean, and live animals crammed together in stalls (Woodward, 2020; Zhu et al., 2020).
References
Center for Infectious Disease Dynamics (CIDD). (2014a, March 19). Week 1 Video 5: Reproductive Number [Video]. https://youtu.be/ju26rvzfFg4
Center for Infectious Disease Dynamics (CIDD). (2014b, March 19). Week 1 Video 6: Epidemic Curve [Video]. https://youtu.be/sSLfrSSmJZM
Cintrón-Arias, A. (2015, November 4). East Tennessee State University MATH 5880 Basic reproductive number [Video]. https://youtu.be/ItW-Q6Npapo
Delamater, P. L., Street, E. J., Leslie, T. F., Yang, Y. T., & Jacobsen, K. H. (2019). Complexity of the basic reproduction number (R0). Emerging infectious diseases, 25(1).
Read, J. M., Bridgen, J. R., Cummings, D. A., Ho, A., & Jewell, C. P. (2020). Novel coronavirus 2019-nCoV: early estimation of epidemiological parameters and epidemic predictions. medRxiv.
Woodward, A. (2020, January 31). The outbreaks of both the Wuhan coronavirus and SARS likely started in Chinese wet markets. Business Insider. https://www.businessinsider.com/wuhan-coronavirus-chinese-wet-market-photos-2020-1
Zhu, H., Guo, Q., Li, M., Wang, C., Fang, Z., Wang, P., … & Xiao, Y. (2020). Host and infectivity prediction of Wuhan 2019 novel coronavirus using deep learning algorithm. bioRxiv.